语言

 中洪博元,动物实验,肺部模型
中洪博元,动物实验,肺部模型
 中国实验动物信息网
中国实验动物信息网
 2020-07-23
2020-07-23
 4149
4149
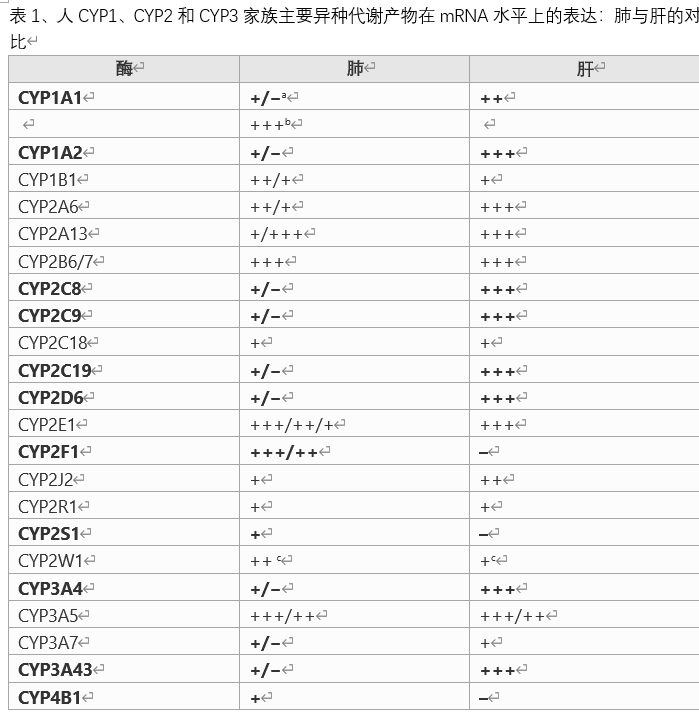
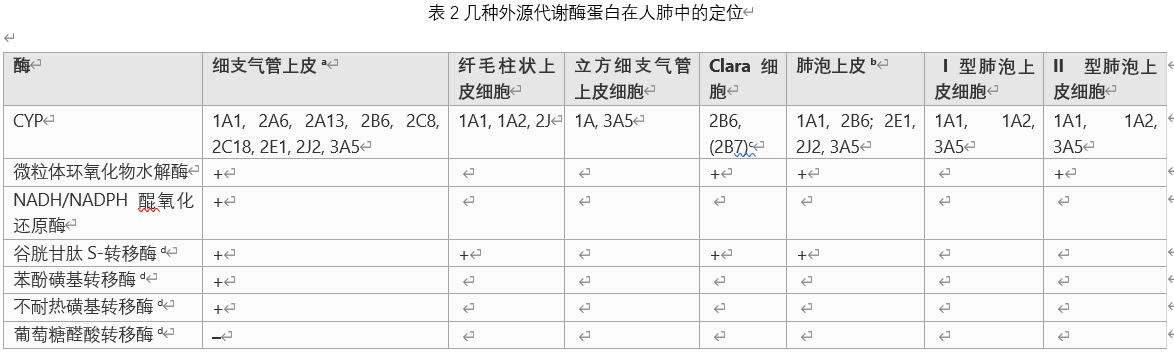
审查从该审查收集的数据以及该审查结束时表11的摘要中,我们可以看到,适当的系统的选择是不同的,只要所考虑的化合物的代谢关键涉及不同的酶即可。 由于化合物的不同性质可能取决于不同代谢物或母体化合物的去除程度,因此即使是特定化合物也可能需要不同的系统来研究不同的性质。哺乳动物的肺是高度复杂的器官,具有许多隔室和40多种不同的细胞类型。本文收集的数据表明,这些不同的细胞区室和细胞类型中外源性代谢酶的模式是不同的。当前可用于识别单个实验动物物种或人类体外模型在模拟人的肺部状况方面的相对适合性的大多数信息是基于整个器官的肺部,例如肺匀浆和部分肺。从...获取这些酶在癌变过程中和在癌细胞中的水平和/或活性的变化,以及多态性及其对癌症易感性的影响均不包括在本综述中,并且可能会成为将来的综述主题。
(1)肺中的某些肺部代谢酶,例如细胞色素P450(CYP),活性很低,难以准确测量。
(2)本评论和/或表格中引用的大部分活动均来自原始出版物,该出版物未说明在一定的线性条件下是否已正确测试了给定的速率。因此,并非所有提供的值都可以进行比较。因此,明智的做法是将本评论中引用的值仅代表高,中,低或未观察到的活动。在大多数“零活动”情况下,作者没有给出检测极限,因此类似的考虑适用于低活动性和非活动性,并且实际上由一位作者给出的低活动性数量是可能低于其他作者给出的值。没有活动。外源代谢酶家族/超家族在首次出现时进行了简要介绍。大鼠肺外源代谢酶:细胞色素P450:细胞色素P450是哺乳动物肝脏和某些肝外组织中主要的外源代谢氧化还原酶。
CYP还原酶:Zhu等。 (1985)在大鼠肺动脉内皮细胞和平滑肌细胞,支气管上皮细胞和平滑肌细胞,I型上皮细胞和巨噬细胞中发现了CYP4A mRNA。 Hellmold(1993)等人报道了CYP4A2和CYP4A8在大鼠肺中的表达。 Zeldin等。 (1996)在大鼠肺中发现了CYP2J3 mRNA。湖等。 (2003)通过在精确切开的大鼠肺切片中暴露于BNF(β-萘黄酮),aroclor1254或BP诱导CYP1A1 mRNA(高达8.3倍)。盖特等。 (2006)报道,大鼠暴露于沥青烟中后,大鼠肺中的CYP1A1和1B1显着增加(26-2203倍),CYP2F2 mRNA水平显着降低(2.4倍)。做到了。非CYP氧化还原酶黄素依赖性单加氧酶(FMO):FMO发生在哺乳动物肝脏中,似乎是肝外组织中一种重要的异源代谢氧化还原酶,后者比CYP更重要可能比这更重要。然而,它们的底物特异性限于软亲核中心中的氮和硫原子,特别是异种生物分子,转化为N-氧化物和S-氧化物。 Sukumaran(2011)等。报告了大鼠肺中FMO2和FMO3 mRNA的存在及其昼夜节律变化。AD(P)H:醌氧化还原酶(NQO):NQO通过直接两电子还原避免半醌自由基作为其主要代谢产物,从而防止了醌毒性。QO将羰基官能团转化为羟基官能团,通常用作葡萄糖醛酸转移酶和/或磺基转移酶结合的底物,以促进排泄。ADH或NADPH可以用作还原当量。 Gate(2006)等人报道了暴露于沥青烟的大鼠肺中NQO1 mRNA的显着增加(6.7倍)。酒精脱氢酶(ADH):ADH将酒精转化为醛或酮。乙醇主要通过ADH转化为乙醛,这会引起乙醇的急性毒性。AD用作电子受体。醛脱氢酶(ALDH):ALDH将醛转化为相应的羧酸。此步骤通常代表排毒。醛氧化酶(AO):AO通过同时形成过氧化氢来催化醛转化为羧酸。使用分子氧作为电子受体。谷胱甘肽S-转移酶(GST):GST将亲电子异物转化为谷胱甘肽(GSH)结合物。它通常是排毒的,但在某些情况下是有毒的。葡萄糖基转移酶:UGT将具有适当取代基(主要是羟基,有时是氨基,巯基或羧基)的外源化合物转化为葡萄糖醛酸。这通常是排毒,但在某些情况下会引起毒性(例如“酰基转移”)。兔肺外源性代谢酶:细胞色素P450:兔肺组织中CYP4B1 mRNA的表达显着高于肝脏和其他组织。
CYP和相关蛋白的表达:Serabjit Singh等。 (1980)观察到CYP还原酶蛋白在兔肺克拉拉细胞中。 CYP还原酶在兔肺克拉细胞中的定位高于II型肺泡巨噬细胞。 CYP还原酶蛋白在兔I型肺细胞和内皮细胞中均无表达。尽管在兔肺组织中未检测到CYP1A1和CYP1A2蛋白,但TCDD可以诱导CYP1A1和CYP1A2蛋白在支气管,支气管上皮,肺动脉,静脉和毛细血管内皮细胞中表达。 CYP1A1在兔肺泡巨噬细胞中含量较低。 CYP2B4在兔子的克拉拉细胞和II型肺细胞,支气管和支气管上皮以及I型肺细胞中观察到。
CYP2B4的相对含量在Clara细胞中较高,而CYP2B4的相对含量在支气管和细支气管纤毛上皮细胞中较低。 CYP4B1在Clara细胞,支气管纤毛细胞,支气管纤毛细胞,II型肺细胞和毛细血管内皮细胞中含量*高,但在II型肺细胞中则不是。非CYP氧化还原酶:黄素依赖性单加氧酶(FMO),醛酮还原酶(AKR)。水解酶:环氧水解酶(EH):微粒体环氧水解酶(mEH,也称为EPHX1),可溶性环氧水解酶(sEH,也称为EPHX2)。金仓鼠肺外源代谢酶:细胞色素P450(CYP):Lorenz等人(1984)表明,金仓鼠肺微粒体可有效催化广谱CYP底物7-乙氧基香豆素。像大鼠肺微粒体一样,乙基比小鼠肺微粒体慢约5倍,但比人肺微粒体快100倍。长期以来,金仓鼠中的肺Clara(Club)细胞已被确定为CYP依赖性共价结合的主要位置。 Paolini等。 (1995)报道了CYP2B依赖的PROD活性在金黄仓鼠肺中和几种化合物(苯巴比妥钠,巴比妥钠,环磷酰胺)的诱导作用。结合酶:谷胱甘肽S-转移酶(GST),磺基转移酶(SULT)。豚鼠肺中的外源性代谢酶:细胞色素P450(CYP):Bilimia等。 (1977年)观察到豚鼠肺总匀浆AHH活性。动物接触香烟烟雾后,AHH活性显着降低(约50%)。猪肺外源代谢酶:细胞色素P450(CYP):在猪肺中观察到CYP1A1和CYP1A2转录本。猪CYP2B22(与人CYP2B6同源,核苷酸同源性为81%)mRNA的组成型表达水平高于猪肝。 CYP2D25(与人CYP2D6同源,核苷酸同源性为83%)在猪肺中低水平表达。猪肺组织含有CYP3A22、3A29和3A46 mRNA,但它们的表达水平远低于肝脏中的表达水平。偶联酶:谷胱甘肽S-转移酶(GST)。狗肺中的外在代谢酶:细胞色素P450(CYP):Visser等。 (2017)观察到CYP2B11转录本在犬肺中的表达以及其他外源代谢酶的mRNA的*低表达。 CYP催化的几种外源性化合物的单加氧反应已在犬肺中观察到。水解酶:环氧水解酶(EH)。偶联酶:谷胱甘肽S-转移酶(GST)。猴肺外源代谢酶:mar猴:细胞色素P450(CYP):Uehara等人(2018年)观察到mar猴肺中高表达的CYP2F1转录本,远超过其他任何研究所。高(肝),肾,空肠,脑)。
免疫印迹显示,mar猴肺CYP2F1蛋白与人CYP2F1抗体交叉反应。在mar猴肺微粒体中,观察到了几种原型CYP底物的催化活性,如香豆素,7-乙氧基香豆素和2-/4-联苯,但未发现氯唑沙宗。食蟹猴:细胞色素P450(CYP):Uehara等人(2018)证实了食蟹猴肺中CYP2F1转录本的高表达。在食蟹猴肺微粒体中,观察到某些典型的CYP底物(如香豆素,7-乙氧基香豆素和2-/4-联苯)的催化活性,但未观察到毒死x。